Archives
- 2026-07
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
The development of novel approaches to study GPCR
2020-05-22
The development of novel approaches to study GPCR-mediated transactivation in live UNC 2400 is important, and this study describes a unique BRET-based quantification of Grb2 recruitment to the EGFR as a direct readout of GPCR-mediated transactivation. We have previously used the recruitment of Grb2
-
br Limitations There are some limitations to
2020-05-22
Limitations There are some limitations to take into account when interpreting these results. Firstly, direct causation cannot be established in this cross-sectional study, and it may be the downstream effects of TNFα, such as increased Mdivi 1 australia production, which are more proximally rela
-
Cys LT have been shown to
2020-05-22
Cys-LT have been shown to be critical for stimulation of monocytes to produce various proinflammatory factors [32,67]. Fig. 3 shows results obtained with human peripheral monocyte-derived macrophages differentiated for 6 days with 20 ng/ml macrophage colony-stimulating factor (M-CSF) and challenged
-
Recent data obtained with etifoxine suggest that facilitatio
2020-05-22
Recent data obtained with etifoxine suggest that facilitation of GABAergic inhibition may be also associated with an indirect mechanism implying activation of the peripheral benzodiazepine receptor with a subsequent increase in neurosteroid production (Schlichter et al., 2000, Verleye et al., 2005).
-
br Discussion The differences in
2020-05-21
Discussion The differences in the kinetic properties of L-AmDH and LeuDH presented herein reflect a different kinetic mechanism. When compared to its parent enzyme, L-AmDH displayed a different substrate binding order, lower affinity for the keto substrate, and stronger product inhibition. The tw
-
br Conflict of interest br Acknowledgements This
2020-05-21
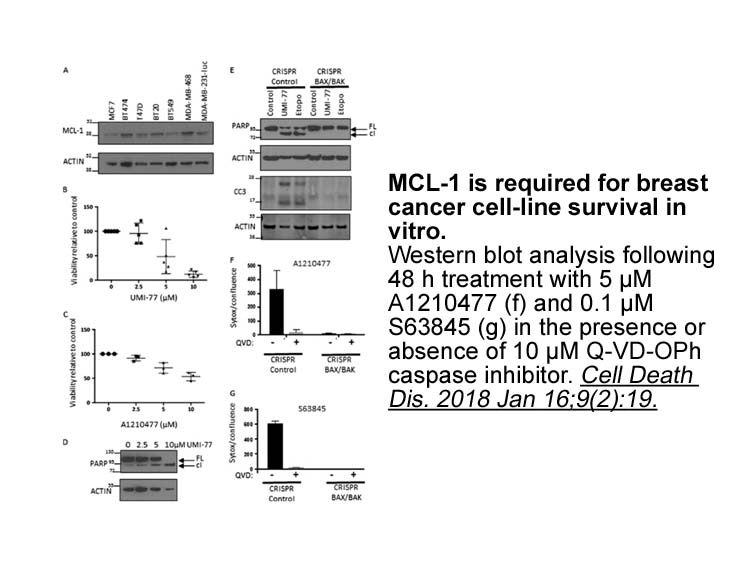
Conflict of interest Acknowledgements This work was supported by the Biotechnology Research Center, China Three Gorges University, Yichang, China (No. 2016KBC05), the National Natural Science Foundation of China (No. 31370600 and 31300508), the Zhejiang Science and Technology Major Program on
-
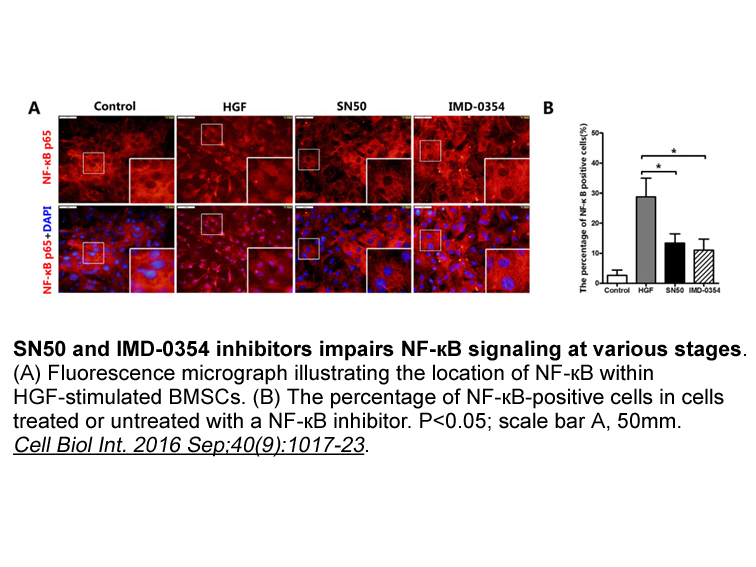
Tumor associated macrophages TAMs one of the limiting factor
2020-05-21
Tumor-associated macrophages (TAMs)—one of the limiting factors in anti-tumor immunity—are associated with the poor clinical outcome in most carcinomas, owing to their potential to promote angiogenesis and local invasion, increase the metastasis of tumor cells, and inhibit the anti-tumor immunity.11
-
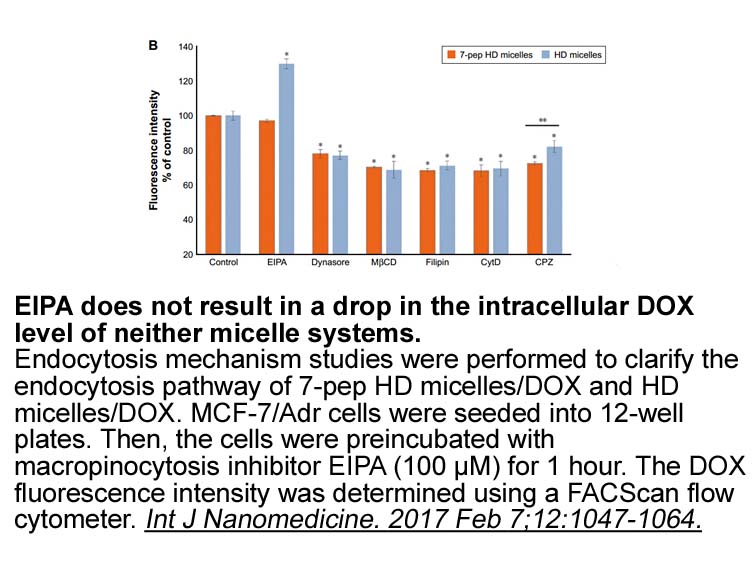
Many authors have described the
2020-05-21
Many authors have described the existence of correlation between viability and moisture content and therefore, water availability. Sabuquillo et al. (2010) demonstrated that only Penicillium oxalicum conidial formulations whose initial moisture contents ranged between 5 and 14% retained their viabil
-
Secondary and primary studies have
2020-05-21
Secondary and primary studies have considered the decision to select the provider and to select components after the component origin is chosen. Examples of secondary and primary studies outside our scope, but still related to decision-making are listed in Tables 1 and 2. The secondary and primary s
-
br COTSget based Architectures This section describes how ou
2020-05-21
COTSget-based Architectures This section describes how our COTSget-based architectures are defined. We use a design inspired by MDE to build a Domain-Specific Language (DSL) of the architecture, as can be seen in the metamodel of Fig. 3. This metamodel defines the abstract syntax of our DSL and i
-
br Materials and methods br
2020-05-21
Materials and methods Results and discussion Iron (II)/2-oxoglutarate (2-OG)-dependent oxygenases [28], [29] catalyze oxidative reactions of a range of metabolic processes including the hydroxylation of l-proline and l-lysine residues during the post-translational modification of collagen. Mor
-
SM-164 CK comprises a family of serine threonine protein
2020-05-21
CK1 comprises a family of serine/threonine protein kinases, that includes different CK1 isoforms namely α, β, δ, ε and γ their various splice variants [56]. It was reported that the pharmacological inhibitor IC261 shows an order of magnitude of higher selectivity for CK1δ or CK1ε isoforms (IC50 of 1
-
br Materials and methods br Results and
2020-05-21
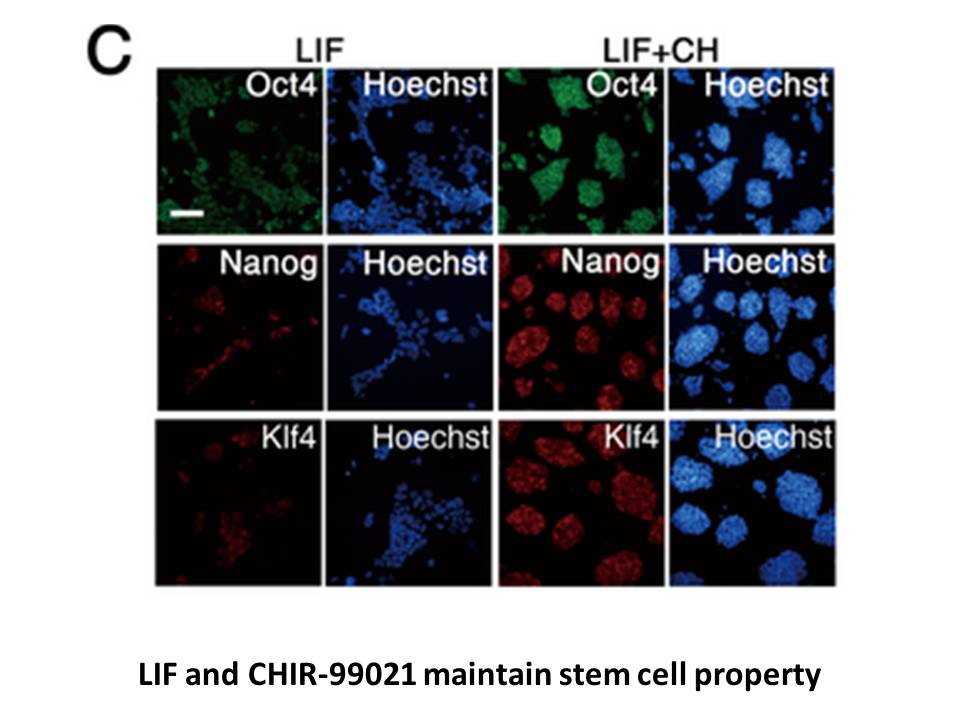
Materials and methods Results and discussion Conclusions In the present study, we have shown the important role of miR-204-5p in C2C12 myoblast differentiation. Overexpression of miR-204-5p inhibited myogenic differentiation and decreased the formation of slow-twitch myofibers, whereas miR-
-
As the first allotetraploid fish in the world
2020-05-21
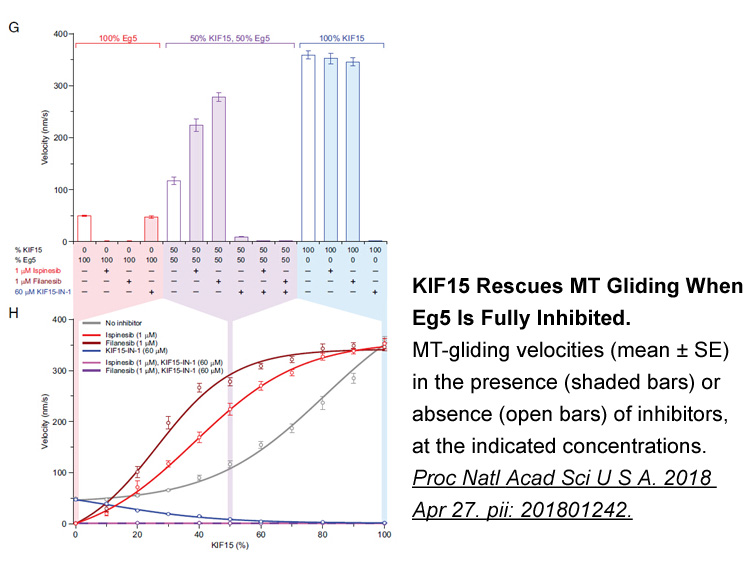
As the first allotetraploid fish in the world, our lab selectively breed the allotetraploid hybrid fish (4nAT, 4n = 200) from the distant hybridization offspring F3 of female Red Crucian Carp (Carassius auratus red var., abbreviated as RCC, 2n = 100) and male Common Carp (Cyprinius carpio L., abbrev
-
During the past three decades platinum
2020-05-21
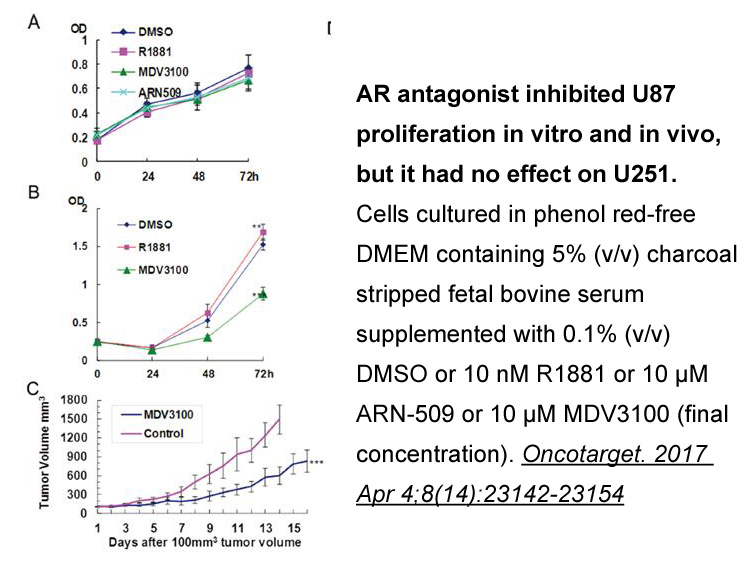
During the past three decades, platinum-based DNA-alkylating agents have been employed in the treatment of various cancers [14]. Oxaliplatin is currently used as a first-line treatment for colorectal cancer (CRC) [15]. Unfortunately, a large portion of CRC patients receiving oxaliplatin-based chemot
16342 records 878/1090 page Previous Next First page 上5页 876877878879880 下5页 Last page